Induced proximity for targeted protein degradation
In 1993, Spencer et al. used the antibiotic rapamycin to bring two proteins into proximity, initiating signal transduction.1 Stuart Schreiber formally described this spatial reorganization of protein-protein interactions as “induced proximity” in 2021; this can be used to categorize post-translational modifications, transcriptional regulation, and targeted degradation.2
The discovery of targeted protein degradation as a deliberate therapeutic strategy is credited to the early 2000s, particularly with the invention of “PROTACs” (PROteolysis TArgeting Chimeras).3 This was the first practical demonstration of disease-causing proteins being selectively marked for degradation via chemically induced proximity to E3 ubiquitin ligases, thereby co-opting the cell’s natural protein degradation systems.
In essence, targeted protein degradation uses small molecules to induce proximity between a protein of interest and an E3 ubiquitin ligase, leading to ubiquitination and subsequent degradation. This approach serves as a promising solution for previously undruggable targets, such as transcription factors and scaffolding proteins. In fact, several lead candidates have been approved, or are in clinical trials, notably for cancer and neurodegenerative diseases.4 The interactions of the degrader small molecules with their targets, however, must be explored in detail to finely tune their behavior and therapeutic potential.
An emerging approach for the visualization of the ternary complexes formed during targeted protein degradation is single particle analysis, a cryogenic electron microscopy (cryo-EM) technique that can produce near-native-state 3D reconstructions of protein complexes. This high-resolution information can reveal the selectivity and cooperativity of complexes and guide the structure-based drug design of novel scaffolds for better degradation efficiency and specificity.
The structural basis of cancer-mutation-driven neomorphic protein-protein interactions
Cancer mutations can create neomorphic protein-protein interactions that drive aberrant function. For instance, as a substrate receptor of the CULLIN3-RING E3 ubiquitin ligase complex, KBTBD4 is recurrently mutated in medulloblastoma (the most common type of embryonal brain tumor). These mutations mediate gain-of-function to KBTBD4, inducing aberrant degradation of CoREST corepressor complexes, which include LSD1 and HDAC1/2.5
Structural analysis via cryo-EM reveals that KBTBD4 mutants form asymmetric dimers, with inserted amino acid residues physically occupying HDAC1’s active site. UM171, a known hematopoietic stem cell agonist, acts as a molecular glue and induces a nearly identical molecular configuration between wild-type KBTBD4 and HDAC1, effectively mimicking the behavior of the cancer mutations.6
These insights provide a rational basis for the development of HDAC1/2 inhibitors via cryo-EM as targeted therapies for cancer-harboring KBTBD4 mutations, and they offer a structural template for the de novo drug discovery of molecular glues.

Targeted protein degradation involved in RNA processing and stress granule formation
CRBN (Cereblon) is a substrate receptor protein within the CRL4/CRBN E3 ubiquitin ligase complex that guides this complex to specific protein targets for ubiquitination and subsequent degradation.
One prominent molecular degrader, lenalidomide (available commercially as REVLIMID), targets CRBN to recruit non-native targets (called neo-substrates) for degradation. Normally, CRBN binds specific proteins via degrons (i.e., recognition motifs such as the G-loop), but molecular glue degraders expand the substrates accessible to CRBN by modifying its surface.
Recently, Monte Rosa Therapeutics discovered MRT-5702, a molecular glue degrader targeting Ras–GAP SH3 domain binding protein 2 (G3BP2). This novel CRBN neo-substrate challenges the idea that the G-loop is the primary binding site, as MRT-5702 induces G3BP2 degradation via molecular surface mimicry; CRBN appears to mimic an endogenous partner of G3BP2’s NTF2L domain.
A high-resolution cryo-EM structure of the CRBN-MRT-5702-G3BP2 ternary complex shows a novel binding pose involving a flexible LON-loop of CRBN, rather than the conventional binding site on its CULT domain.7 This observation could guide the design of next-generation molecular-glue degraders that target different protein-protein interaction hotspots on disease-relevant proteins.

Development of heterobifunctional degraders for cancer immunotherapy
Enzymes, such as phosphatases, are challenging drug targets because they lack a classical ATP binding pocket and their active sites are highly polar; this makes them an attractive target for targeted protein degradation.
Protein tyrosine phosphatase 2 (PTPN2), and the closely related PTPN1, are tumor proteins that could be targeted with this approach to increase immunotherapy efficacy. A first-in-class small-molecule active-site inhibitor was recently discovered that targets these enzymes and displays potent anti-tumor activity; it is currently in phase 1 clinical trials for the treatment of locally advanced or metastatic tumors.8
PTPN2/N1 hetero-bifunctional degraders (Cmpd-1 and Cmpd-2) were also found and form efficient complexes with the CRL4-CRBN E3 ligase complex; these have been shown to mediate potent target degradation in cells and in vivo. Both compounds form cooperative ternary complexes with CRBN–DDB1 and selectively degrade PTPN2/N1. Cmpd-1 shows superior potency in vitro and in vivo. A cryo-EM structure reveals ternary complex formation of PTPN2–Cmpd-1–CRBN/DDB1, highlighting dynamic conformations and weak direct contacts between PTPN2 and CRBN.
This study showcases the importance of cryo-EM in accessing dynamic, non-static interactions in ternary complex formation, which is essential for the rational design of targeted degraders. It also suggests that structural plasticity, rather than rigid binding, may underly effective ubiquitination and degradation, demonstrating a viable targeted protein degradation strategy for phosphatases using CRBN-recruiting heterobifunctional degraders.

Addressing undruggable targets in the human proteome with targeted protein degradation
The integration of novel degrader scaffolds and cryo-EM structural analysis is redefining drug discovery for the undruggable human proteome. Since many degrader ternary complexes are large, flexible, and difficult to crystallize, cryo-EM is highly suitable for the high-resolution structural analysis of these complexes in their native state and within dynamic assemblies. It enables the rational design of degraders with improved selectivity, cooperativity, and biological outcomes, paving the way for next-generation therapeutics across oncology, neurodegeneration, and immunology.
References
- Spencer DM, et al. Controlling Signal Transduction with Synthetic Ligands. Science 262:5139 (1993). doi: 10.1126/science.7694365
- Schreiber SL. The Rise of Molecular Glues. Cell 184:1 (2021). doi: 10.1016/j.cell.2020.12.020
- Sakamoto KM, et al. Protacs: Chimeric molecules that target proteins to the Skp1–Cullin–F box complex for ubiquitination and degradation. Proc Natl Acad Sci USA 98:15 (2001). doi: 10.1073/pnas.141230798
- Hu Y, et al. Molecular glue degrader for tumor treatment. Frontiers 14 (2024). doi: 10.3389/fonc.2024.1512666
- Xie X, et al. Converging mechanism of UM171 and KBTBD4 neomorphic cancer mutations. Nature 639 (2025). doi: 10.1038/s41586-024-08533-3
- Yeo MJR, et al. UM171 glues asymmetric CRL3–HDAC1/2 assembly to degrade CoREST corepressors. Nature 639 (2025). doi: 10.1038/s41586-024-08532-4
- Annunziato S, et al. Molecular surface mimicry enables CRBN to target G3BP2 for degradation. bioRxiv 04.30.651496 (2025). doi: 10.1101/2025.04.30.651496
- Hao Q, et al. Mechanistic insights into a heterobifunctional degrader-induced PTPN2/N1 complex. Commun Chem 7:183 (2024). doi: 10.1038/s42004-024-01263-7

Advances in high-resolution cryo-EM at 100 kV
100 kV cryo-TEM enables high-resolution single particle anal... Alex Ilitchev, PhD
Read More
3D Tissue Histology with Light-Sheet Microscopy Enables Nondestructive Analysis of Microglia
3D tissue analysis offers critical benefits for neuroscience... Alex Ilitchev, PhD
Read More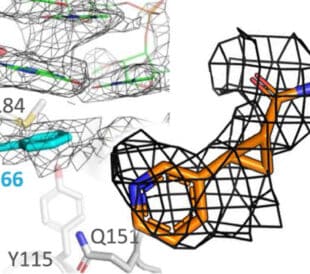
Fragment based drug discovery meets challenging drug targets with high-throughput cryo-EM
Benefits of FBDD in the search for novel therapeutics Frag... Dominic Meusch
Read More
Analyzing Microparticles for Drug Delivery: the Critical Role of SEM and FIB-SEM Technology
Microparticles in drug development and delivery Microparticl...
Read More
Leave a Reply